

Mismatch negativity (MMN) is a neurophysiological response measured as an auditory event-related potential (ERP) generated automatically by the brain when a deviant sound occurs rarely and unpredictably amongst regular background sounds. A reduction in the amplitude of MMN in schizophrenia is one of the most replicated results in schizophrenia studies first reported approximately 30 years ago. Considering the large effect size of the MMN reduction and other characteristics of MMN in schizophrenia, it has been proposed that MMN have the status of a neurophysiological biomarker of the disorder. In this paper, we review the evidence that has accrued to support the assertion of biomarker status, and then evaluate what that means for diagnosis, prognosis, and prediction of medication response, but most importantly, its use as a translational tool in pre-clinical studies. Although considerable knowledge has accrued over the last decades about the optimal stimulus conditions for exposing MMN impairments in schizophrenia, the association with cognitive deficits and functional status, and the involvement of the glutamate NMDAr system, little is known about the neurobiological basis of MMN reduction in schizophrenia. Pre-clinical research using animal models is essential to make advances in this area. Fortunately, paradigms have been developed for use in the rat that strongly suggest that the rat brain is capable of generating the equivalent of a human MMN response or MMR. We review evidence supporting that a late component of MMR in the rat exhibits many of the characteristics of the human MMR. What is needed now is an animal model of schizophrenia that produces a robust and replicable reduction in this component that can be investigated using molecular approaches to support the development of novel pharmaceuticals to rescue MMR in these animals.
2)School of Biomedical Science and Pharmacy, College of Health, Medicine and Wellbeing, University of Newcastle
3)Centre for Brian and Mental Health Research, University of Newcastle
4)Hunter Medical Research Institute
Introduction
The first publication reporting that the mismatch negativity (MMN) component of the auditory event-related potential was reduced in schizophrenia was by an Australian group in 199182), followed shortly afterwards by a similar report from a New York group39)in 1993. In the intervening 30 years, MMN amplitude reduction was noted in over 500 separate reports (Web of Science count excluding reviews) in patients with a diagnosis of a schizophrenia spectrum disorder from a variety of laboratories and patient sources (hospitalized, outpatients, supported accommodation, etc. ). Numerous meta-analyses estimated the effect size (EF) to be of the order of 0.8-1.0 depending on the exact methodology used to generate the response14)97). The robustness of this finding along with other attributes of MMN in schizophrenia have led to assertions that reduced MMN meets the criteria for a biomarker of the disorder4)55). The purpose of this review is to summarize the evidence that has accrued to support the assertion of biomarker status, and then evaluate what that means for diagnosis, prognosis, or prediction of medication response99), but most importantly, its use as a translational tool in pre-clinical studies78). First, a brief description of MMN, the stimulus conditions that generate a mismatch response, and models of mechanisms proposed as explanations of the MMN signal are provided.
What is MMN?
MMN was first described by a Finnish researcher, Risto Näätänen, and his colleagues in the late 70s60)61)in the context of a program of research to understand the effects of selective attention on event-related potentials (ERPs) of the brain. MMN is an electrophysiological response to rare deviant or oddball stimuli that interrupt regularity in background standard stimulation (Figure 1A). It has been observed in auditory, visual, and somatosensory modalities, but is more frequently examined in audition, mainly because of ease of stimulus control. MMN can be readily recorded with scalp electrodes (Figure 1C37)) using standard electroencephalography (EEG) systems. There is a magnetoencephalography (MEG) equivalent-the MMNm. It is an automatic response to stimulus change-that is, its occurrence is not reliant on active attention being paid to the attributes of the eliciting stimulus sequence. This attribute of automaticity is one of the reasons why it has been such a useful tool for investigating brain function in clinical patients with cognitive impairments. For auditory stimuli, MMN consists of a negative potential of approximately 1-4μV peaking at approximately 150-250 ms after the deviant event, and is most often visualized and measured by subtracting the ERP response to the regular standard sounds from the ERP response to deviant sounds (Figure 1B). MMN is elicited by any deviant that represents a discriminable violation of a regularity in regular sounds, such as a change in a simple attribute〔e. g. frequency (pitch), duration, spatial location〕, or a violation of a more complex regularity such as a pattern violation (e. g. regular tone pairs of low-high pitch interrupted by a deviant pair of high-low where the absolute frequencies of sounds making up the pair are randomized77)), speech sounds, or stimulus omission in a repeating sound sequence64). MMN is generally maximal (most negative) at frontocentral sites and reverses in polarity over mastoid and more posterior sites when recorded with a nose reference (Figure 1D). There are at least two sources of activity contributing to the scalp potential-a bilateral supratemporal source contributing to the polarity reversed potential recorded over temporal and posterior sites and a right hemisphere frontal source in general, the degree of laterality depending upon the nature of the eliciting stimulus, with language stimuli producing a more left lateralized response64).
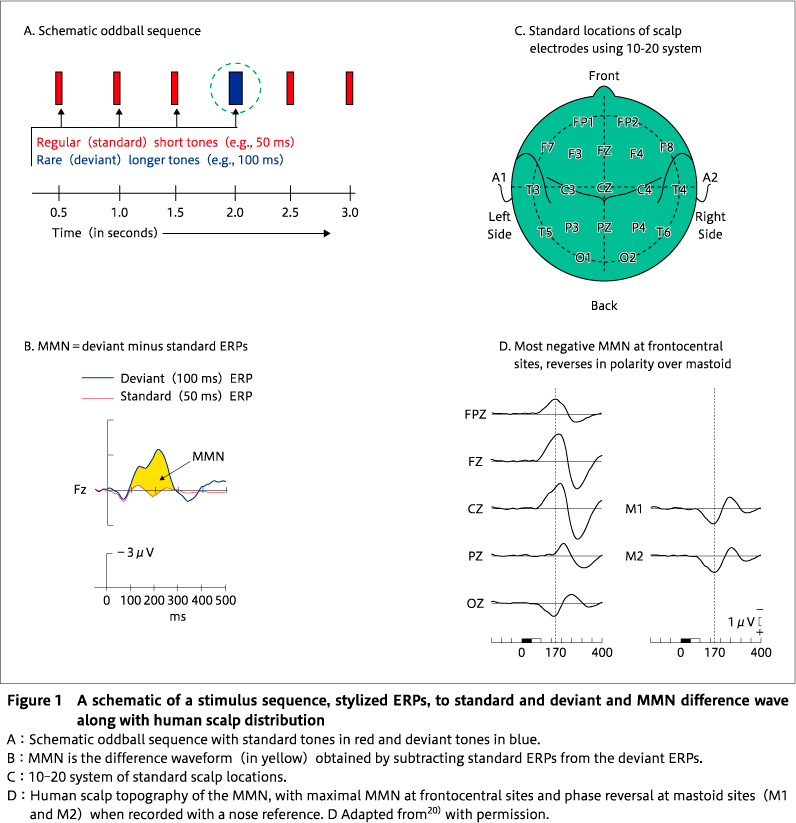
Neuronal mechanisms of MMN
Several models of the neuronal mechanisms underpinning the generation of the MMN have been proposed. The classic interpretation of the MMN proposes that a sensory memory (or echoic memory) of acoustic regularities stored in the auditory cortex supports the detection of sensory deviance between the memory and sensory input62)64). Fronto-temporal networks are then engaged to assess the relevance of the deviance and trigger an attention switch towards the change15). It is assumed that the sum of the neural activity supporting these processes is reflected in the recorded scalp MMN and its topography100). However, this model was challenged by the proposal that the MMN waveform is generated as a result of different levels of neuronal adaptation of an exogenous auditory ERP component, namely the N1 component, generated in response to a novel sound (the deviant) versus a repeated sound (the standard) 34). This model denied the necessity for any special sensory memory or genuine deviance detection, but was firmly rebutted on a variety of grounds63), but most importantly that MMN can be elicited even in the absence of an N1 (as in the stimulus omission paradigm101)) or repetition of the same sounds77). However, both explanations of MMN generation can be integrated under a single unifying framework of a predictive coding model21). Predictive coding is a general theory of brain function underpinning perceptual inference and learning19)that proposes that the brain is a hierarchically organized system with feedforward and feedback connections between levels of the system that attempt to minimize discrepancies between bottom-up sensory inputs provided from a lower level, such as auditory areas located in temporal regions, and top-down predictions from a higher level (frontal areas) 21). According to this framework, a novel sound that violates predictions (based on an environmental model of past regularities) about what the next stimulus will be, generates a prediction error necessitating an updated environmental model, reflected in MMN. A repeated sound that matches predictions results in a suppression of prediction errors through a process akin to adaptation. Hence, the predictive coding account incorporates both adaptation mechanisms and a model (or memory) of past regularities. Furthermore, the predictive coding framework emphasizes the importance of short-term plasticity of glutamatergic synaptic connections between lower and higher brain regions that underpins model updating, the latter regulated by glutamate N-methyl-D-aspartate receptors (NMDARs) 81). Numerous pharmacological studies in humans and animals revealed that the MMN amplitude is reduced by the administration of NMDAR antagonists40)95)94).
MMN as a biomarker for a diagnosis of schizophrenia
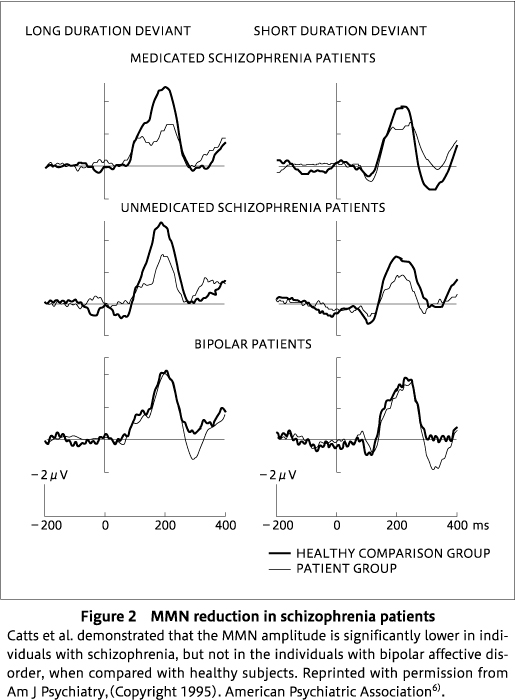
As noted earlier, reduced MMN in medicated patients with a diagnosis of schizophrenia was first observed in the early 1990s using simple paradigms in which the deviant was a variation either in duration82)or frequency(pitch)39). Shortly afterwards, the Australian group reported that the reduced MMN to a duration deviant also occurred in unmedicated patients with a schizophrenia diagnosis, but was absent in patients with a bipolar diagnosis6), demonstrating not only that reduced MMN occurred irrespective of medication status in schizophrenia patients, but also that the effect was restricted to patients with a diagnosis of schizophrenia(Figure 2), suggesting specificity of the marker to schizophrenia rather than a general marker of psychosis(see also96) for a similar result, although meta-analysis14)suggests a small but significant reduction of MMN in bipolar patients). There is some evidence that a duration deviant is a more sensitive dimension than frequency based on the same group of patients in whom frequency MMN was unaffected, whereas duration MMN was substantially reduced in amplitude57). The first meta-analysis of 62 studies of MMN in schizophrenia offered some support for the notion that MMN to duration deviants was more impaired in schizophrenia than that to frequency deviants, although it failed to reach significance with an effect size of 1.01 for duration and 0.47 for frequency97). On the other hand, a subsequent meta-analysis of an even larger set of 104 unique studies found evidence of a significantly larger MMN deficit to duration than frequency deviants14). However, the real situation may be more complex, as we reported that reduction of MMN to duration deviants is evident even in the early stages of the illness, whereas the reduction of MMN to frequency deviants occurs in later stages92). We57)previously suggested that extraction of the duration dimension of sound requires more complex computational processing within multiple auditory networks31)than frequency, which is robustly coded in multiple tonotopically mapped regions of the auditory cortex86). Thus, duration may be more vulnerable to even subtle anatomical or connectivity changes that occur early in the course of schizophrenia.
Considering the impressive effect size of 1.0, particularly for a duration deviant, in distinguishing between a healthy control group and a group with a diagnosis of schizophrenia, how useful is MMN likely to be as a diagnostic marker? Admittedly, such an effect size means that, on average, 84% of a patient group will have an MMN amplitude lower than the control group, but there are still many impediments to MMN amplitude being used as a diagnostic biomarker or to move it into clinical use56)80). Studies reporting MMN data in schizophrenia populations vary substantially in terms of their methods:physical stimulus parameters〔frequency(Hz), duration, probability of deviant, etc.〕, recording system parameters(filter settings, digitization rate, and amplification), recording montage(location of active sites and reference location), and post-processing algorithms(artefact rejection criteria)80). This lack of standardization and normative data, including age norms, combined with lack of extensive data on sensitivity and specificity to schizophrenia are a major impediment to using MMN as a diagnostic biomarker.
There are, however, other attributes of MMN reduction that strengthen the case for it being designated as a diagnostic biomarker of schizophrenia. The MMN measure has utility for investigating the cognitive aspects of auditory processing65), a link that is strengthened by reframing MMN within a predictive coding framework of brain functioning21). Impaired cognition is a central feature of schizophrenia and is largely responsible for the poor functional outcomes of patients in work, education, and relationships leading to high-levels of life long disability22)23). Although the number of published MMN studies in schizophrenia that examine correlations between MMN and cognitive performance is limited(for a review93)), a large multi-center study of 1415 patients found, using structural equation modelling, that an early auditory processing measure driven largely by reduced MMN has a direct effect on cognition. Poor functioning is also associated with reduced MMN in patients with functioning assessed using a variety of scales, including the Global Assessment of Function(GAF)53)54)and Social and Occupational Assessment of Functioning(SOFAS)73). The large multi-center study cited earlier89)found that MMN amplitude predicted functional outcomes and cognition, the authors estimating that an intervention that resulted in a 1-μV increase in MMN amplitude will be theoretically linked to improvements in both cognition and functional outcome(effect sizes of 0.78 and 0.28, respectively). Therefore, the development of interventions that increase MMN amplitude is an important goal for clinical researchers.
NMDAR deficiency has been implicated in the cognitive deficits and symptoms of schizophrenia. A direct comparison of the effects of ketamine, an NMDAR antagonist, and amphetamine, a DA agonist, administered to healthy volunteers revealed that only ketamine produced cognitive deficits and negative symptoms that mimic those exhibited by schizophrenia patients50). This and other evidence challenged the dominant model of schizophrenia as being due to a dopaminergic imbalance. Ketamine was also demonstrated to reduce MMN in healthy volunteers94). Furthermore, the amplitude of MMN prior to ingestion predicted the level of psychotic experiences induced by ketamine95). MMN therefore indexes the degree to which the NMDAr system is vulnerable to disruption by NMDAr antagonists and may reflect the functional state of the glutamate NMDAr neurotransmission. Thus, reduced MMN in schizophrenia has been touted as a marker of aberrant glutamatergic neurotransmission67).
MMN as a prognostic biomarker
Individuals can be at-risk of developing schizophrenia either on the basis of genetic risk(having a first-degree relative with the disorder)or on the basis of sub-threshold clinical symptoms(low-level symptoms below the threshold for a diagnosis of a psychotic disorder accompanied by a decline in social or occupational functioning), referred to as a clinically defined high-risk group. There is mixed evidence on whether first-degree relatives exhibit reduced MMN with early evidence suggesting that MMN is as reduced in first-degree relatives as in diagnosed patients58), but the most recent meta-analysis14)based on 8 separate reports suggested that MMN in this group is quantitatively(EF=0.26), but not significantly, reduced. Evidence for reduced MMN in clinically defined high-risk individuals however is stronger, with meta-analysis14)producing a modest and significant reduction relative to healthy controls(EF=0.40).
However, is there evidence that MMN size can predict who will develop psychosis? The first report5)that this may be the case used a Cox regression model to demonstrate that MMN to duration deviants(but not frequency deviants)predicted who will develop psychosis, mostly with a schizophrenia diagnosis. A later report71)demonstrated that the size of the MMN elicited by a double deviant differing in duration and pitch from standards not only predicted who will develop, but also the time from ERP assessment to psychosis onset in clinically at-risk individuals. In summary, there is evidence that the degree of MMN amplitude reduction in at-risk individuals is useful as a prognostic biomarker in predicting those with low-level psychotic symptoms who will develop a schizophrenic disorder, but the same deficiencies that limit the usefulness of MMN as a diagnostic biomarker also apply to MMN as a prognostic marker, i. e., the lack of standardization of methods and normative data.
MMN as a biomarker of treatment outcome
It has been argued that one of the most promising areas for investment in schizophrenia biomarkers is in predicting treatment response-not only existing therapies, but most importantly for developing novel therapies99). There are a number of characteristics of MMN that are relevant to the issue of treatment effects. First, does MMN in patients respond to current neuroleptic therapies? The answer is in general no. Neither first nor second generation neuroleptics ameliorate MMN deficits in schizophrenia patients. The one exception is aripiprazole, a relatively new antipsychotic with a unique receptor binding profile that combines partial agonistic activity at D2 receptors and 5-HT 1A receptors, and potent antagonism at 5-HT 2A receptors104). After 4 weeks of treatment with aripiprazole, MMN amplitudes to both frequency and duration deviants improved with further improvement after 8 weeks104). In addition, increased MMN was accompanied by reduced positive, negative and total PANSS ratings, and correlated with decreased total PANSS ratings, suggesting that MMN improvement is a possible biomarker of treatment efficacy104). These results contrast with the lack of effects of other atypical antipsychotics, such as clozapine, risperidone, and olanzapine, on MMN(reviewed in104)).
Second, is there evidence that novel treatments are effective in increasing MMN in patients? There have been reports of heightened MMN with novel pharmacotherapies that target glutamatergic neurotransmission. Administration of the glutathione precursor N-acetyl-cysteine(NAC)as adjunctive therapy to standard neuroleptic medication significantly increased MMN in schizophrenia patients after 6 weeks of treatment51). In addition, glycine, which functions as a co-agonist with glutamate of NMDA receptor activity in excitatory glutamatergic neurotransmission, again administered as adjunctive therapy, increased MMN acutely in patients, but not after 6 weeks of treatment24). Similar acute effects of a high dose of memantine, a moderate affinity NMDAr antagonist, delivered as adjunctive therapy, significantly increased MMN in patients, although the degree of enhancement was smaller in patients than in healthy subjects85). Although the memantine effects appear paradoxical considering its NMDAr antagonism, it has been used as adjunctive therapy in schizophrenia to improve cognition, although with limited success46).
Third, is there any evidence that MMN can predict the clinical response to current therapies? This important question is infrequently investigated. A relatively old but preliminary report79)on a three-year follow-up of clinical response to clozapine treatment in patients found that MMN amplitude measured prior to clozapine therapy commencement is related to treatment response, although perhaps not in the direction expected:a relatively intact MMN was associated with a better prognosis79). The interpretation of this result is that a good clinical response to clozapine is associated with relatively intact brain functioning as indicated by a larger MMN. Although unexpected, it nonetheless suggests that MMN has value in clinical decisions about medication choice.
Can MMN itself be used to develop novel pharmacotherapies? Although reduced MMN in schizophrenia was first observed approximately 30 years ago, it has not led to improved pharmacotherapies for patients, primarily because human studies are limited in terms of probing the biological mechanisms of disrupted MMN generation. Hence, the need for translational studies of MMN reduction in animal models, which is the focus of the remainder of this review.
MMN as a translational biomarker in schizophrenia
Evaluation of translational biomarkers in schizophrenia assumes that a valid animal model of this complex heterogeneous psychiatric disorder can be developed. The fact that schizophrenia is a uniquely human disorder has complicated the development of suitable animal models. Animal homologues of positive and negative symptoms, such as hallucinations, delusions, alogia, or affective flattening, are clearly problematic with behavioral assessments utilized as animal equivalents of positive45)and negative12)symptoms requiring further validation. In recent years, there has been a greater focus on developing animal models that mimic the cognitive deficits that patients exhibit. However, this approach also has limitations because of difficulties in mapping particular human cognitive constructs. For example, working memory is usually assessed in humans by archetypal tasks such as digit span backwards or the n-back task. In animal models, considering the constraints imposed by species differences in terms of shaping(instructing)and measuring behavior, there is uncertainty about whether the selected animal task targets the same construct in the animal and human. The advantage of using an electrophysiological measure, such as MMN, is that it can be measured in exactly the same manner in animals and humans using the same stimulus sequences, and the same methodology for recording and processing data. There is still an issue of determining whether the animal brain can produce the equivalent of an adaptation-independent mismatch response(MMR)that reflects prediction error. This issue is discussed below. First however, we address the question of what is the primary impetus for moving into animal models of this complex neuropsychiatric disorder.
1.Why do we need animal models of schizophrenia?
Although animal model studies provide evidence for what is changed, and how it is changed in schizophrenia, they can more importantly shed light on mechanisms of how symptoms or behaviors are induced, and how such outcomes can be ameliorated or treated. For example, although the glutamatergic hypothesis of schizophrenia was triggered by the clinical observations that NMDAr antagonists, such as phencyclidine(PCP), or ketamine can induce a syndrome that resembles schizophrenia in humans10)42)49)50)93), it was animal model studies that provided evidence for the mechanisms underlying these effects using pharmacological and genetic approaches33)52)72). Another example of the significance of animal models is demonstrated by the treatment studies performed with compounds acting directly at the NMDAR glycine site, such as glycine, D-serine, or D-cycloserine, and, more recently, with high-affinity glycine transport inhibitors42). A significant improvement in schizophrenia-like negative symptoms in a NMDAR antagonist rat model using dizocilpine(MK-801)was demonstrated in the presence of D-serine or by blocking glycine transporter-1 that stimulates the NMDAr function directly or indirectly, respectively44). Administration of D-cycloserine, a co-agonist at the glycine site on the NMDAr, attenuated attentional deficits in rats treated with MK-80132). However, although the current gold standard method for evaluating the face validity of "schizophrenia-ness" of a given animal model is behavioral, behavioral measurements are limited in their ability to translate animal model outcomes to humans18). Therefore, recent attention has turned to searching for biomarkers of schizophrenia that can be measured in a similar manner in rodents and humans.
2.Animal models of MMN
Due to clinical relevance of the MMN for evaluating normal and pathological auditory processing, the search for evidence of MMN in several mammalian species, including cats8), guinea pigs48), monkeys38), rats76), and mice98), is underway in order to facilitate further understanding of the neurophysiological and neurochemical mechanisms underlying MMN generation, and for exploring the mechanism that leads to MMN reduction. Among other species, rodents are well-suited for preclinical studies due to their small size, easy handling, and faster breeding cycles, which makes them cost-efficient models. In general, rats are often the preferred species for behavioral and neuroscience research because of their relatively larger brains, but mice are more suited to molecular studies13). Initial studies observed MMN-like ERPs 63-253 ms post stimulus onset, recorded epidurally from the auditory cortex of anesthetized rats76). In addition, dependence of MMN generation on the presence of standard tones in a sequence was found in anesthetized rats using DEV-alone condition, where human-like MMN was not observed in response to DEV tones in the DEV-alone paradigm vs an oddball paradigm, consistent with human studies demonstrating the dependence of human MMN on the echoic trace76). In another study on urethane-anesthetized rats, an MMN-like response was observed at 60-100 ms in response to ascending DEV from stimulus onset during an auditory oddball sequence2). Several studies demonstrated MMN-like responses in anesthetized rats1-3) and awake rats27)30)68)74)84). Overall, there is strong evidence to support the presence of MMN-like responses in both anesthetized and awake rats.
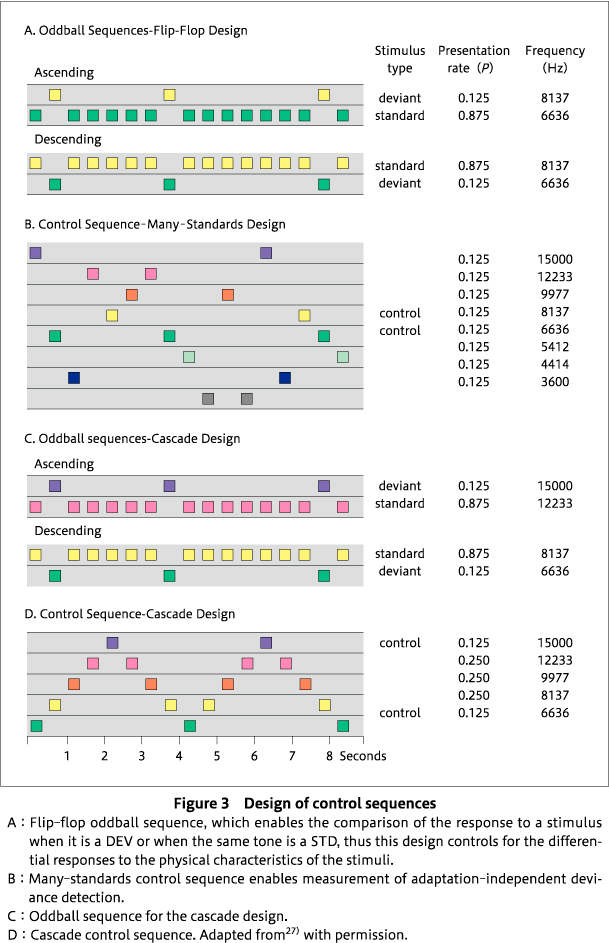
A methodological feature making MMN a valuable candidate for translation into animals is that the MMN in humans is elicited without instructing the participant to attend to the stimuli. Similarly, the recording of mismatch responses (MMR) in animals does not require the active participation of the animal (no prior training is required), provided the animal can discriminate the sounds in the sequences that are used to generate MMR. Second, the auditory patterns that elicit MMN can be simple, for example, auditory stimuli of different tone frequencies, as frequency discrimination thresholds have been well established in rodents87). However, the face validity of an animal model of MMN needs to be established: how does a rat MMR resemble human MMN? For preclinical research, it is essential to show the adaptation-independent element of human MMN in rodents. Harms and colleagues provided evidence that the rat brain is capable of generating human-like MMN, at least in terms of having some element of adaptation-independent prediction error (a criteria of human MMN) 29)68), by employing three different control sequences-1. the flip-flop control (to control for differential responses to the physical characteristics of standards and deviants as in35)); 2. the many-standards control (to control for differential adaptation as in35)), and 3. the cascade control (to control for differential adaptation) 27). In the flip-flop oddball sequences, two oddball sequences are present such that the identity of the standard and deviant reverse, which means that a given tone frequency appears as standard in one sequence and a deviant in the other sequence (Figure 3A). Use of the flip-flop sequences enables the control of the physical characteristics of the deviant and standard stimuli. In the many-standards control sequence, the equivalent of the deviant tone from the oddball sequence is presented among other equiprobable tones equal to the probability of the deviant in the oddball sequence and the tones are presented pseudo-randomly such that no pattern of regularity is established (Figure 3B). Thus, no prediction can be set due to lack of regularity and no violation can occur. A comparison of the responses to the deviant in the oddball sequence to the same sound at the same probability of occurrence in the many-standards control sequence excludes the explanation that lower levels of adaptation of neural populations responding to the deviant are responsible for the larger response to a deviant stimulus. Instead, this comparison yields a measure of adaptation-independent deviance detection or prediction error27)69). However, the many-standards control has been subjected to two criticisms. First, as the many-standards sequence contains stimuli from a broad frequency range, larger responses are produced than with those from a narrower range, as the deviant response from oddball undergoes more adaptation (thus, reduced size amplitude) than the deviant response to the many-standards control, which is not affected much by adaptation (thus, increased in amplitude). Overall, this potential imbalance may result in an underestimation of deviance detection. Second, the oddball sequence establishes a repetitive and predictable sequence that is violated, but the many-standards control, by design, does not violate a prediction because one is never established.
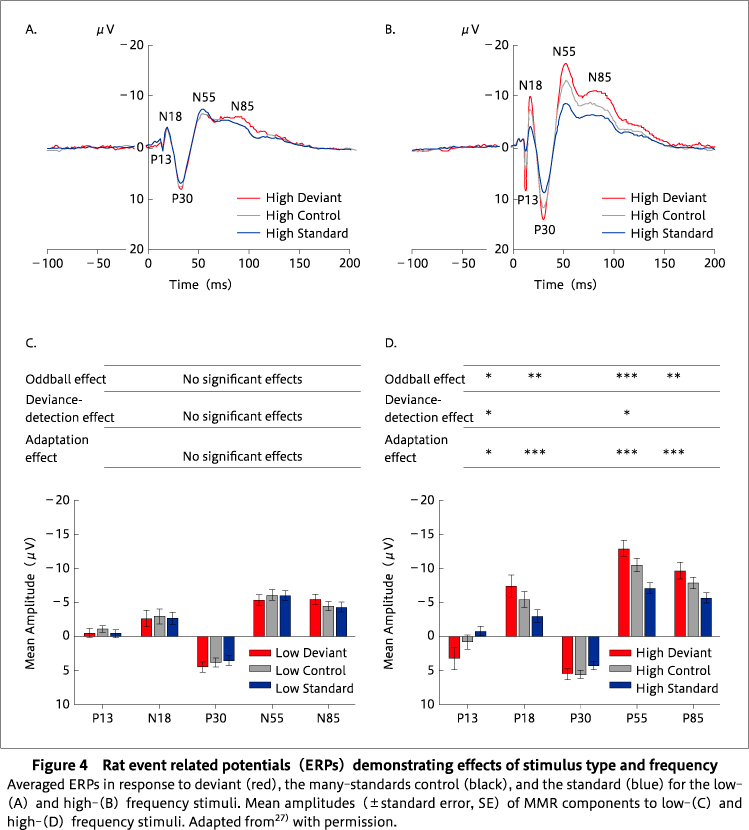
To overcome these issues, the cascade control, initially proposed by Schroger's group75), was used by Harms and colleagues. In the cascade control, stimuli are presented varying from low to high frequency in a regular pattern, with the highest frequency stimulus corresponding to the deviant and the second highest frequency stimulus corresponding to the standard within an ascending oddball sequence (Figure 3C, D) 27). Unexpectedly, there were inconclusive results in the cascade control design, possibly because rats may be unable to encode the complex pattern of the ascending and descending arms of the cascade control sequence. However, other researchers recording single unit and multiple unit activity in oddball paradigms from auditory brain regions found no difference in outcomes between the cascade control and the many-standards control69)70), although only ascending or descending cascade sequences were used. Therefore, adaptation-independent deviance detection was only evident for the many-standards control sequence, not for the cascade sequence in27) (Figure 4). As shown in Figure 4A and 4B, larger ERPs are generated in response to high frequency deviants, but not low frequency deviants, suggesting the lack of effect on descending deviants27). Studies conducted in our laboratory68)and other groups7)26)43)83)84)demonstrated that rodents exhibit an adaptation-independent MMR, similar to human MMN, which can even be observed in local recordings from multiple auditory processing regions69). Of note, it has only recently been demonstrated that this adaptation-independent component of human MMN is reduced in patients with schizophrenia47).
3.Do animal mismatch responses(MMRs)exhibit features of human MMN?
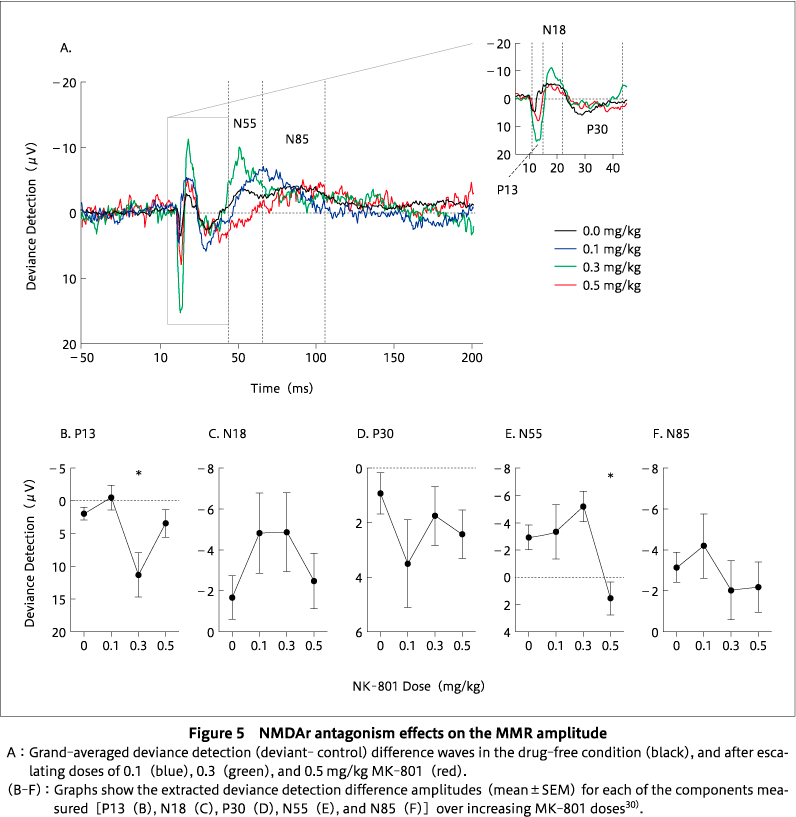
To establish the face validity of an animal model of MMN, there are simple requirements, the first being that a deviant response> standard response. To date, animal studies have reported this, as described above, although not for low-frequency deviants in our laboratory. Some of the key features of human MMN are adaptation independence (as described above), sensitivity to the NMDAR perturbation, sensitivity to deviance difference, sensitivity to deviant probability, and sensitivity to environmental uncertainty (`jitter'). In order to establish the face validity of an animal model of MMN, there is a need to determine how well the animal model exhibits these features. Studies have demonstrated consistent reductions in the MMN amplitude in humans in the presence of NMDAr antagonists such as ketamine and phencyclidine25)41)42)88)93). Human MMN amplitude increases in response to the higher degree of deviance difference between the standard and the deviant stimuli, lower probability of occurrence of the deviant, and a stable stimulus onset asynchrony (SOA) in between the two auditory stimuli (reviewed in36)). Several studies reported that NMDAr hypofunction also reduces MMRs in rodents11)17)28)30)70)84)90)91). We found that the hypothesized NMDAr antagonism exclusively affects the N55 MMR component. There was a reduction in the size of MMR amplitude for late negative potential (N55) at the highest dose of the antagonist, but not on an early positive potential (P13) (Figure 5). This suggests that N55 is the most MMN-like component30). However, there is only limited evidence of the sensitivity of the rodent MMR to the degree of deviance, DEV probability, and environmental uncertainty.
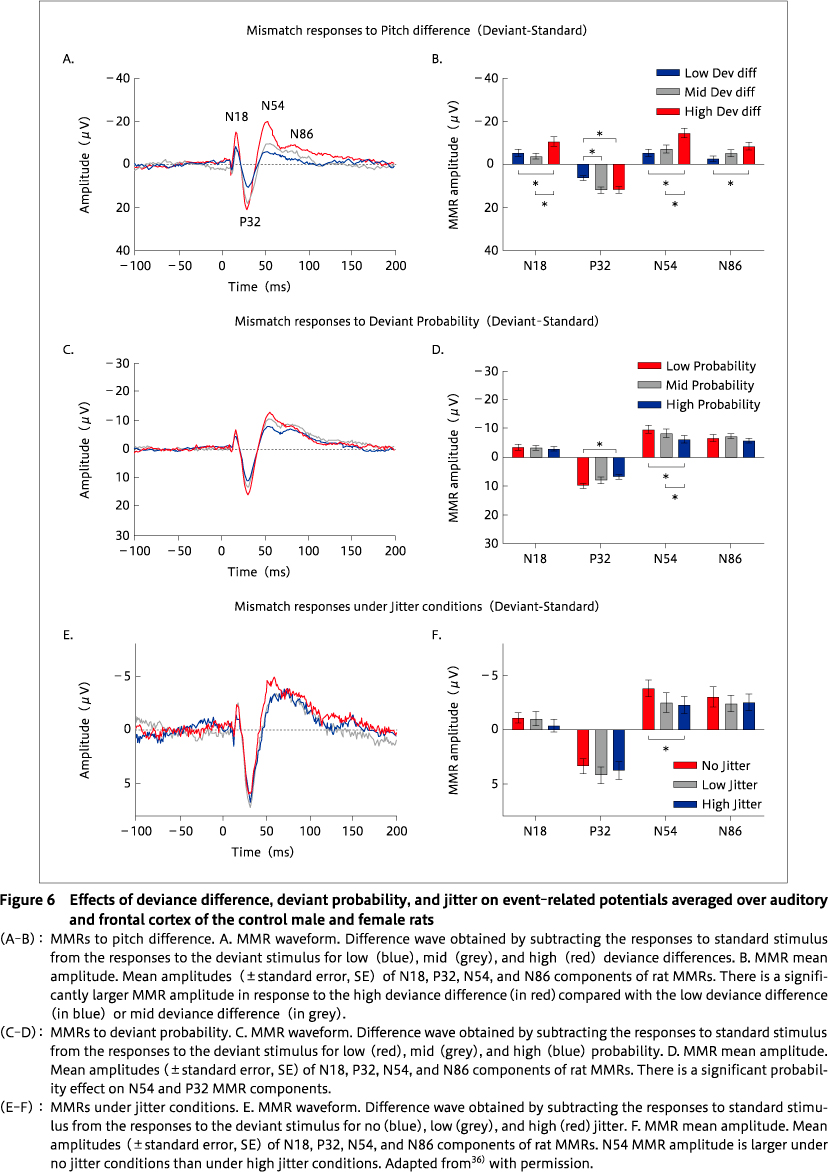
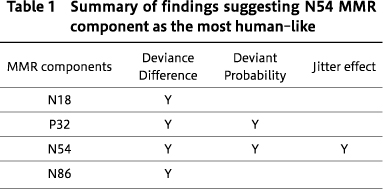
Considering the growing research interest in the rodent MMRs as a translatable`biomarker'for schizophrenia, it is important to examine how reliably rat MMRs mimic the attributes of human MMN. An extensive review on how well the MMN features recapitulate human features in the studies of rodent MMRs is presented in36). Although there are increasing preclinical studies on MMRs, some gaps in understanding remain. To fill some of these gaps, our laboratory used a rat model of MMN to determine the degree to which the rat brain can recapitulate features of MMN found in healthy human controls. We asked whether MMRs in rats exhibit features of human MMN, such as larger MMR amplitude, in response to 1. a higher degree of deviance difference between the regular standard tones and surprising deviant tones, 2. lower probability of occurrence of deviant tone among regular standards, and 3. stable SOA (no jitter). Screw electrodes were surgically-implanted epidurally, and using wireless telemetry in freely-moving rats, we observed human-like alterations of MMRs36). Rat MMRs decreased in size with a reduced degree of difference between the deviant and standard stimuli, with increased probability of the deviant stimuli, and with increased variability in the SOA of sound sequences (Figure 6). This suggested that N54 MMR component in rats is highly`MMN-like', as it was significantly influenced by all 3 alterations (Table 1) 36). The strongest effects of stimulus type were previously observed in response to high-frequency stimuli for the N55 component of MMR27). Furthermore, Harms et al. noted MK-801-induced N55 MMR amplitude reduction, similar to human MMN reductions, elicited by NMDAr antagonism30). The N54 component by Jalewa et al. 36)is equivalent to the N55 component reported by Harms et al. 30). The different labelling is the result of a naming convention of using the mean peak latency across all animals in a study. Overall, these studies suggest that rodent MMR mimics human MMN attributes and that N54/N55 is the most-human like MMN analogue in rats.
4. What are the implications of animal MMRs for schizophrenia?
Although MMN impairment in schizophrenia patients is one of the most highly replicated neurophysiological features of the disorder66), there are few studies demonstrating that MMN reduction is well-treated by current antipsychotic drugs24)104). Compounds that pass the pre-clinical stages in developing novel psychiatric treatments often fail in human clinical trials due to lack of suitable animal models of disease with associated biomarkers of neural-circuit function across species55)102)103). Moreover, examining MMRs in rodents is a fairly new field of research, and has only been pursued for the past 10 years or so, with relatively few studies having looked for MMRs in rats or mice, model species for which a wealth of existing knowledge, techniques, and relevant methodologies is available17)90). Examination of the clinical literature reviewed earlier demonstrates that there is much that we still do not know about MMN in schizophrenia. Animal models are required in order to advance our understanding of the neuro-circuitry of MMN generation and to provide knowledge about the neurobiological basis of MMN reduction in schizophrenia59). This knowledge is essential for the development of novel pharmaceuticals that can rescue the impairments in function that underlie the reduced amplitude of MMN. As noted earlier, a 1-μV increase in MMN amplitude will likely result in improvements in both cognition and functional outcome89).
Conclusion
There are several approaches to creating a rodent model of MMN reduction in schizophrenia, including acute pharmacological (NMDAR antagonists) challenge11)30)40)90)and genetic manipulations16), and endogenous causes such as exposure to neurodevelopmental insults associated with schizophrenia risk9). The purpose of such approaches is not to recapitulate schizophrenia in an animal model, but to explore the details (the what, why, and how) of the underlying neurobiological mechanisms that result in attenuation of MMN amplitude. Although we have learnt much from acute pharmacological challenges based on NMDAR antagonism, it is likely that a model based on endogenous causes producing a large, enduring, and reproducible reduction of MMN will be more informative. It may enable multiple approaches, including molecular approaches, to exploring the causes of MMN reduction, and the development of new pharmaceuticals targeting increased MMN based on this knowledge. Pre-clinical research of this type may also offer insights into the optimal paradigms for use in humans-are there particular features of the oddball paradigm that are more likely to generate a reduced MMN in animal models and why?
[Acknowledgements]
JJ is supported by a University of Newcastle Postgraduate Research Scholarship funded by the Australian Government's Research Training Program. Clinical research was funded by the Australian National Health and Medical Research through project grants #880840, #900663, #960579, and #990701. Pre-clinical research was funded by the Australian National Health and Medical Research grants ID 1026070 and ID 1109283.
[Disclosure Statement]
None of the co-authors have conflicts of interest with any companies or organizations that may influence the study reported here.
[Author Contributions]
PM and JJ wrote the manuscript. JJ prepared Figure 1. All authors reviewed the content, and contributed to and approved the final manuscript.
1) Astikainen, P., Ruusuvirta, T., Wikgren, J., et al.: Memory-based detection of rare sound feature combinations in anesthetized rats. Neuroreport, 17 (14); 1561-1564, 2006![]()
2) Astikainen, P., Stefanics, G., Nokia, M., et al.: Memory-based mismatch response to frequency changes in rats. PLoS One, 6 (9); e24208, 2011![]()
3) Astikainen, P., Mällo, T., Ruusuvirta, T., et al.: Electrophysiological evidence for change detection in speech sound patterns by anesthetized rats. Front Neurosci, 8; 374, 2014![]()
4) Belger, A., Yucel, G. H., Donkers, F. C.: In search of psychosis biomarkers in high-risk populations: is the mismatch negativity the one we've been waiting for? Biol Psychiatry, 71 (2); 94-95, 2012![]()
5) Bodatsch, M., Ruhrmann, S., Wagner, M., et al.: Prediction of psychosis by mismatch negativity. Biol Psychiatry, 69 (10); 959-966, 2011![]()
6) Catts, S. V., Shelly, A. M., Ward, P. B., et al.: Brain potential evidence for an auditory sensory memory deficit in schizophrenia. Am J Psychiatry, 152 (2); 213-219, 1995![]()
7) Chen, I. W., Helmchen, F., Lütcke, H.: Specific early and late oddball-evoked responses in excitatory and inhibitory neurons of mouse auditory cortex. J Neurosci, 35 (36); 12560-12573, 2015![]()
8) Csepe, V., Karmos, G., Molnar, M.: Evoked potential correlates of stimulus deviance during wakefulness and sleep in cat: animal model of mismatch negativity. Electroencephalogr Clin Neurophysiol, 66 (6); 571-578, 1987![]()
9) Dunn, A. L., Michie, P. T., Hodgson, D. M., et al.: Adolescent cannabinoid exposure interacts with other risk factors in schizophrenia: a review of the evidence from animal models. Neurosci Biobehav Rev, 116; 202-220, 2020![]()
10) Egerton, A. A., Grace, A., Stone, J., et al.: Glutamate in schizophrenia: neurodevelopmental perspectives and drug development. Schizophr Res, 223; 59-70, 2020![]()
11) Ehrlichman, R. S., Maxwell, C. R., Majumdar, S., et al.: Deviance-elicited changes in event-related potentials are attenuated by ketamine in mice. J Cogn Neurosci, 20 (8); 1403-1414, 2008![]()
12) Ellenbroek, B. A., Cools, A. R.: Animal models for the negative symptoms of schizophrenia. Behav Pharmacol, 11 (3-4); 223-233, 2000![]()
13) Ellenbroek, B., Youn, J.: Rodent models in neuroscience research: is it a rat race? Dis Model Mech, 9 (10); 1079-1087, 2016![]()
14) Erickson, M. A., Ruffle, A., Gold, J. M.: A meta-analysis of mismatch negativity in schizophrenia: from clinical risk to disease specificity and progression. Biol Psychiatry, 79 (12); 980-987, 2016![]()
15) Escera, C., Alho, K., Winkler, I., et al.: Neural mechanisms of involuntary attention to acoustic novelty and change. J Cogn Neurosci, 10 (5); 590-604, 1998![]()
16) Featherstone, R. E., Shin, R., Kogan, J. H., et al.: Mice with subtle reduction of NMDA NR1 receptor subunit expression have a selective decrease in mismatch negativity:implications for schizophrenia prodromal population. Neurobiol Dis, 73; 289-295, 2015![]()
17) Featherstone, R. E., Melnychenko, O., Siegel, S. J.: Mismatch negativity in preclinical models of schizophrenia. Schizophr Res, 191; 35-42, 2018![]()
18) Forrest, A. D., Coto, C. A., Siegel, S. J.: Animal models of psychosis:current state and future directions. Curr Behav Neurosci Rep, 1 (2); 100-116, 2014![]()
19) Friston, K.: Learning and inference in the brain. Neural Netw, 16 (9); 1325-1352, 2003![]()
20) Fulham, W. R., Michie, P. T., Ward, P. B., et al.: Mismatch negativity in recent-onset and chronic schizophrenia: a current source density analysis. PLoS One, 9 (6); e100221, 2014![]()
21) Garrido, M. I., Kilner, J. M., Stephan, K. E., et al.: The mismatch negativity: a review of underlying mechanisms. Clin Neurophysiol, 120 (3); 453-463, 2009![]()
22) Green, M. F.: What are the functional consequences of neurocognitive deficits in schizophrenia? Am J Psychiatry, 153 (3); 321-330, 1996![]()
23) Green, M. F.: Cognitive impairment and functional outcome in schizophrenia and bipolar disorder. J Clin Psychiatry, 67 (10); e12, 2006![]()
24) Greenwood, L. M., Leung, S., Michie, P. T., et al.: The effects of glycine on auditory mismatch negativity in schizophrenia. Schizophr Res, 191; 61-69, 2018![]()
25) Gunduz-Bruce, H., Reinhart, R. M. G. , Roach, B. J., et al.: Glutamatergic modulation of auditory information processing in the human brain. Biol Psychiatry, 71 (11); 969-977, 2012![]()
26) Hamm, J. P., Yuste, R.: Somatostatin interneurons control a key component of mismatch negativity in mouse visual cortex. Cell Rep, 16 (3); 597-604, 2016![]()
27) Harms, L., Fulham, W. R., Todd, J., et al.: Mismatch negativity(MMN)in freely-moving rats with several experimental controls. PLoS One, 9 (10); e110892, 2014![]()
28) Harms, L.: Mismatch responses and deviance detection in N-methyl-D-aspartate (NMDA) receptor hypofunction and developmental models of schizophrenia. Biol Psychol, 116; 75-81, 2016![]()
29) Harms, L., Michie, P. T., Näätänen, R.: Criteria for determining whether mismatch responses exist in animal models: focus on rodents. Biol Psychol, 116; 28-35, 2016![]()
30) Harms, L., Fulham, W. R., Todd, J., et al.: Late deviance detection in rats is reduced, while early deviance detection is augmented by the NMDA receptor antagonist MK-801. Schizophr Res, 191; 43-50, 2018![]()
31) He, J.: Long-latency neurons in auditory cortex involved in temporal integration:theoretical analysis of experimental data. Hear Res, 121 (1-2); 147-160, 1998![]()
32) Howe, W. M., Burk, J. A.: Dizocilpine-induced accuracy deficits in a visual signal detection task are not present following D-cycloserine administration in rats. Eur J Pharmacol, 577 (1-3); 87-90, 2007![]()
33) Inta, D., Monyer, H., Sprengel, R., et al.: Mice with genetically altered glutamate receptors as models of schizophrenia:a comprehensive review. Neurosci Biobehav Rev, 34 (3); 285-294, 2010![]()
34) Jääskeläinen, I. P., Ahveninen, J., Bonmassar, G., et al.: Human posterior auditory cortex gates novel sounds to consciousness. Proc Natl Acad Sci U S A, 101 (17); 6809-6814, 2004![]()
35) Jacobsen, T., Schröger, E.: Measuring duration mismatch negativity. Clin Neurophysiol, 114 (6); 1133-1143, 2003![]()
36) Jalewa, J., Todd, J., Michie, P. T., et al.: Do rat auditory event related potentials exhibit human mismatch negativity attributes related to predictive coding? Hear Res, 399; 107992, 2021![]()
37) Jasper, H.: Report of the committee on methods of clinical examination in electroencephalography. Clin Neurophysiol, 112 (4); 713-719, 1958
38) Javitt, D. C., Schroeder, C. E., Steinschneider, M., et al.: Demonstration of mismatch negativity in the monkey. Electroencephalogr Clin Neurophysiol, 83 (1); 87-90, 1992![]()
39) Javitt, D. C., Doneshka, P., Zylberman, I., et al.: Impairment of early cortical processing in schizophrenia:an event-related potential confirmation study. Biol Psychiatry, 33 (7); 513-519, 1993![]()
40) Javitt, D. C., Steinschneider, M., Schroeder, C. E., et al.: Role of cortical N-methyl-D-aspartate receptors in auditory sensory memory and mismatch negativity generation: implications for schizophrenia. Proc Natl Acad Sci U S A, 93 (21); 11962-11967, 1996![]()
41) Javitt, D. C.: Intracortical mechanisms of mismatch negativity dysfunction in schizophrenia. Audiol Neurootol, 5 (3-4); 207-215, 2000![]()
42) Javitt, D. C., Zukin, S. R., Heresco-Levy, U., et al.: Has an angel shown the way? Etiological and therapeutic implications of the PCP/NMDA model of schizophrenia. Schizophr Bull, 38 (5); 958-966, 2012![]()
43) Jung, F., Stephan, K. E., Backes, H., et al.: Mismatch responses in the awake rat:evidence from epidural recordings of auditory cortical fields. PLoS One, 8 (4); e63203, 2013![]()
44) Kawaura, K., Koike, H., Kinoshita, K., et al.: Effects of a glycine transporter-1 inhibitor and D-serine on MK-801-induced immobility in the forced swimming test in rats. Behav Brain Res, 278; 186-192, 2015![]()
45) Kesby, J. P., Eyles, D. W., McGrath, J. J., et al.: Dopamine, psychosis and schizophrenia: the widening gap between basic and clinical neuroscience. Transl Psychiatry, 8 (1); 30, 2018![]()
46) Kishi, T., Ikuta, T., Oya, K., et al.: Anti-dementia drugs for psychopathology and cognitive impairment in schizophrenia: a systematic review and meta-analysis. Int J Neuropsychopharmacol, 21 (8); 748-757, 2018![]()
47) Koshiyama, D., Kirihara, K., Tada, M., et al.: Reduced auditory mismatch negativity reflects impaired deviance detection in schizophrenia. Schizophr Bull, 46 (4); 937-946, 2020![]()
48) Kraus, N., McGee, T., Littman, T., et al.: Nonprimary auditory thalamic representation of acoustic change. J Neurophysiol, 72 (3); 1270-1277, 1994![]()
49) KrystaL, J. H., D'souza, D. C., Mathalon, D., et al.: NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: toward a paradigm shift in medication development. Psychopharmacology (Berl), 169 (3-4); 215-233, 2003![]()
50) Krystal, J. H., Perry, E. B. Jr., Gueorguieva, R., et al.: Comparative and interactive human psychopharmacologic effects of ketamine and amphetamine: implications for glutamatergic and dopaminergic model psychoses and cognitive function. Arch Gen Psychiatry, 62 (9); 985-994, 2005![]()
51) Lavoie, S., Murray, M. M., Deppen, P., et al.: Glutathione precursor, N-acetyl-cysteine, improves mismatch negativity in schizophrenia patients. Neuropsychopharmacology, 33 (9); 2187-2199, 2008![]()
52) Lee, G., Zhou, Y.: NMDAR Hypofunction Animal Models of Schizophrenia. Front Mol Neurosci, 12; 185, 2019![]()
53) Light, G. A., Braff, D. L.: Mismatch negativity deficits are associated with poor functioning in schizophrenia patients. Arch Gen Psychiatry, 62 (2); 127-136, 2005![]()
54) Light, G. A., Braff, D. L.: Stability of mismatch negativity deficits and their relationship to functional impairments in chronic schizophrenia. Am J Psychiatry, 162 (9); 1741-1743, 2005![]()
55) Light, G. A., Näätänen, R.: Mismatch negativity is a breakthrough biomarker for understanding and treating psychotic disorders. Proc Natl Acad Sci U S A, 110 (38); 15175-15176, 2013![]()
56) Light, G. A., Swerdlow, N. R., Thomas, M. L., et al.: Validation of mismatch negativity and P3a for use in multi-site studies of schizophrenia:characterization of demographic, clinical, cognitive, and functional correlates in COGS-2. Schizophr Res, 163 (1-3); 63-72, 2015![]()
57) Michie, P. T., Budd, T. W., Todd, J., et al.: Duration and frequency mismatch negativity in schizophrenia. Clin Neurophysiol, 111 (6); 1054-1065, 2000![]()
58) Michie, P. T., Innes-Brown, H., Todd, J., et al.: Duration mismatch negativity in biological relatives of patients with schizophrenia spectrum disorders. Biol Psychiatry, 52 (7); 749-758, 2002![]()
59) Michie, P. T., Malmierca, M. S., Harms, L., et al.: The neurobiology of MMN and implications for schizophrenia. Biol Psychol, 116; 90-97, 2016![]()
60) Näätänen, R., Gaillard, A. W., Mantysalo, S.: Early selective-attention effect on evoked potential reinterpreted. Acta Psychol (Amst), 42 (4); 313-329, 1978![]()
61) Näätänen, R., Michie, P. T.: Early selective-attention effects on the evoked potential:a critical review and reinterpretation. Biol Psychol, 8 (2); 81-136, 1979![]()
62) Näätänen, R., Alho, K.: Mismatch negativity: a unique measure of sensory processing in audition. Int J Neurosci, 80 (1-4); 317-337, 1995![]()
63) Näätänen, R., Jacobsen, T., Winkler, I.: Memory-based or afferent processes in mismatch negativity (MMN): a review of the evidence. Psychophysiology, 42 (1); 25-32, 2005![]()
64) Näätänen, R., Paaviliainen., P., Rinne, T., et al.: The mismatch negativity (MMN) in basic research of central auditory processing: a review. Clin Neurophysiol, 118 (12); 2544-2590, 2007![]()
65) Näätänen, R., Sussman, E. S., Salisbury, D., et al.: Mismatch negativity (MMN) as an index of cognitive dysfunction. Brain Topogr, 27 (4); 451-466, 2014![]()
66) Näätänen, R., Todd, J., Schall, U.: Mismatch negativity (MMN) as biomarker predicting psychosis in clinically at-risk individuals. Biol Psychol, 116; 36-40, 2016![]()
67) Nagai, T., Kirihara, K., Tada, M., et al.: Reduced Mismatch Negativity is Associated with Increased Plasma Level of Glutamate in First-episode Psychosis. Sci Rep, 7 (1); 2258, 2017![]()
68) Nakamura, T., Michie, P. T., Fulham, W. R., et al.: Epidural auditory event-related potentials in the rat to frequency and duration deviants: evidence of mismatch negativity? Front Psychol, 2; 367, 2011![]()
69) Parras, G. G., Nieto-Diego, J., Carbajal, G. V., et al.: Neurons along the auditory pathway exhibit a hierarchical organization of prediction error. Nat Commun, 8 (1); 2148, 2017![]()
70) Parras, G. G., Valdés-Baizabal, C., Harms, L., et al.: The effect of NMDA-R antagonist, MK-801, on neuronal mismatch along the rat auditory thalamocortical pathway. Sci Rep, 10 (1); 12391, 2020![]()
71) Perez, V. B., Woods, S. W., Roach, B. J., et al.: Automatic auditory processing deficits in schizophrenia and clinical high-risk patients: forecasting psychosis risk with mismatch negativity. Biol Psychiatry, 75 (6); 459-469, 2014![]()
72) Ramsey, A. J.: NR1 knockdown mice as a representative model of the glutamate hypothesis of schizophrenia. Prog Brain Res, 179; 51-58, 2009![]()
73) Rasser, P. E. , Schall, U. , Todd, J. , et al.: Gray matter deficits, mismatch negativity, and outcomes in schizophrenia. Schizophr Bull, 37 (1); 131-140, 2011![]()
74) Roger, C., Hasbroucq, T., Rabat, A., et al.: Neurophysics of temporal discrimination in the rat: a mismatch negativity study. Psychophysiology, 46 (5); 1028-1032, 2009![]()
75) Ruhnau, P., Herrmann, B., Schröger, E.: Finding the right control: the mismatch negativity under investigation. Clin Neurophysiol, 123 (3); 507-512, 2012![]()
76) Ruusuvirta, T., Penttonen, M., Korhonen, T.: Auditory cortical event-related potentials to pitch deviances in rats. Neurosci Lett, 248 (1); 45-48, 1998![]()
77) Saarinen, J., Paavilainen, P., Schöger, E., et al.: Representation of abstract attributes of auditory stimuli in the human brain. Neuroreport, 3 (12); 1149-1151, 1992![]()
78) Sahin, M., Jones, S. R., Sweeney, J. A., et al.: Discovering translational biomarkers in neurodevelopmental disorders. Nat Rev Drug Discov, 18; 235-236, 2019![]()
79) Schall, U., Catts, S. V., Karayanidis, F., et al.: Auditory event-related potential indices of fronto-temporal information processing in schizophrenia syndromes: valid outcome prediction of clozapine therapy in a three-year follow-up. Int J Neuropsychopharmacol, 2 (2); 83-93, 1999![]()
80) Schall, U.: Is it time to move mismatch negativity into the clinic? Biol Psychol, 116; 41-46, 2016![]()
81) Schmidt, A., Diaconescu, A. O., Kometer, M., et al.: Modeling ketamine effects on synaptic plasticity during the mismatch negativity. Cereb Cortex, 23 (10); 2394-2406, 2013![]()
82) Shelley, A. M., Ward, P. B., Catts, S. V., et al.: Mismatch negativity:an index of a preattentive processing deficit in schizophrenia. Biol Psychiatry, 30 (10); 1059-1062, 1991![]()
83) Shiramatsu, T. I., Kanzaki, R., Takahashi, H.: Cortical mapping of mismatch negativity with deviance detection property in rat. PLoS One, 8 (12); e82663, 2013![]()
84) Sivarao, D. V., Chen, P., Yang, Y., et al.: NR2B antagonist CP-101,606 abolishes pitch-mediated deviance detection in awake rats. Front Psychiatry, 5; 96, 2014![]()
85) Swerdlow, N. R., Bhakta, S., Chou, H. H., et al.: Memantine effects on sensorimotor gating and mismatch negativity in patients with chronic psychosis. Neuropsychopharmacology, 41 (2); 419-430, 2016![]()
86) Talavage, T. M., Sereno, M. I., Melcher, J. R., et al.: Tonotopic organization in human auditory cortex revealed by progressions of frequency sensitivity. J Neurophysiol, 91 (3); 1282-1296, 2004![]()
87) Talwar, S. K., Musial, P. G., Gerstein, G. L.: Role of mammalian auditory cortex in the perception of elementary sound properties. J Neurophysiol, 85 (6); 2350-2358, 2001![]()
88) Thiebes, S., Leicht, G., Curic, S., et al.: Glutamatergic deficit and schizophrenia-like negative symptoms: new evidence from ketamine-induced mismatch negativity alterations in healthy male humans. J Psychiatry Neurosci, 42 (4); 273-283, 2017![]()
89) Thomas, M. L., Green, M. F., Hellemann, G., et al.: Modeling deficits from early auditory information processing to psychosocial functioning in schizophrenia. JAMA Psychiatry, 74 (1); 37-46, 2017![]()
90) Tikhonravov, D., Neuvonen, T., Pertovaara, A., et al.: Effects of an NMDA-receptor antagonist MK-801 on an MMN-like response recorded in anesthetized rats. Brain Res, 1203; 97-102, 2008![]()
91) Tikhonravov, D., Neuvonen, T., Pertovaara, A., et al.: Dose-related effects of memantine on a mismatch negativity-like response in anesthetized ratsDose-related effects of memantine on a mismatch negativity-like response in anesthetized rats. Neuroscience, 167 (4); 1175-1182, 2010![]()
92) Todd, J., Michie, P. T., Schall, U., et al.: Deviant matters:duration, frequency, and intensity deviants reveal different patterns of mismatch negativity reduction in early and late schizophrenia. Biol Psychiatry, 63 (1); 58-64, 2008![]()
93) Todd, J., Harms, L., Schall, U., et al.: Mismatch negativity: translating the potential. Front Psychiatry, 4; 171, 2013![]()
94) Umbricht, D., Schmid, L., Koller, R., et al.: Ketamine-induced deficits in auditory and visual context-dependent processing in healthy volunteers:implications for models of cognitive deficits in schizophrenia. Arch Gen Psychiatry, 57 (12); 1139-1147, 2000![]()
95) Umbricht, D., Koller, R., Vollenweider, F. X., et al.: Mismatch negativity predicts psychotic experiences induced by NMDA receptor antagonist in healthy volunteers. Biol Psychiatry, 51 (5); 400-406, 2002![]()
96) Umbricht, D., Koller, R., Schmid, L., et al.: How specific are deficits in mismatch negativity generation to schizophrenia? Biol Psychiatry, 53 (12); 1120-1131, 2003![]()
97) Umbricht, D., Krljes, S.: Mismatch negativity in schizophrenia: a meta-analysis. Schizophr Res, 76 (1); 1-23, 2005![]()
98) Umbricht, D., Vyssotki, D., Latanov, A., et al.: Deviance-related electrophysiological activity in mice: is there mismatch negativity in mice? Clin Neurophysiol, 116 (2); 353-363, 2005![]()
99) Weickert, C. S., Weickert, T. W., Pillai, A., et al.: Biomarkers in schizophrenia: a brief conceptual consideration. Dis Markers, 35 (1); 3-9, 2013![]()
100) Winkler, I., Karmos, G., Näätänen, R.: Adaptive modeling of the unattended acoustic environment reflected in the mismatch negativity event-related potential. Brain Res, 742 (1-2); 239-252, 1996![]()
101) Yabe, H., Tervaniemi, M., Reinikainen, K., et al.: Temporal window of integration revealed by MMN to sound omission. Neuroreport, 8 (8); 1971-1974, 1997![]()
102) Young, J. W., Geyer, M. A.: Developing treatments for cognitive deficits in schizophrenia: the challenge of translation. J Psychopharmacol, 29 (2); 178-196, 2015![]()
103) Young, J. W., Light, G. A.: Cross-species neurophysiological biomarkers of attentional dysfunction in schizophrenia: bridging the translational gap. Neuropsychopharmacology, 43 (1); 230-231, 2018![]()
104) Zhou, Z., Zhu, H., Chen, L.: Effect of aripiprazole on mismatch negativity (MMN) in schizophrenia. PLoS One, 8 (1); e52186, 2013![]()


